Bài viết được tham vấn chuyên môn cùng Bác sĩ cao cấp, Bác sĩ chuyên khoa II Đoàn Dư Đạt - Bác sĩ Nội đa khoa - Khoa khám bệnh & Nội khoa - Bệnh viện Đa khoa Quốc tế Vinmec Hạ Long.
Phản ứng viêm của cơ thể có thể là cấp tính (kéo dài trong vài ngày) còn gọi là phản ứng viêm cấp, hoặc có thể là mãn tính (phản ứng lại sự xúc phạm liên tục và không được giải quyết). Tình trạng viêm có thể phát triển thành tổn thương mô vĩnh viễn hoặc xơ hóa. Trong bài viết này, chúng ta sẽ xem xét các quá trình liên quan đến cơ chế gây phản ứng viêm cấp tính.
Thế nào là phản ứng viêm: Viêm là phản ứng bảo vệ bình thường của cơ thể trước các tổn thương do các tác nhân gây bệnh. Viêm xảy ra khi các tế bào bạch cầu trong cơ thể tăng cường hoạt động để bảo vệ cơ thể khỏi bị đả kích bởi tác nhân gây bệnh.
Các đặc điểm lâm sàng trọng điểm của tình trạng viêm cấp tính (sưng – nóng – đỏ - đau) đã được công nhận ít nhất 5000 năm. Tiến sĩ John Hunter, nổi tiếng vào cuối thế kỷ 18 bác sĩ phẫu thuật người Scotland, quan sát thấy rằng các phản ứng viêm không phải là một căn bệnh mà là một phản ứng không cụ thể và mang tính khởi đầu của sự hoạt động của hệ thống miễn dịch. Thông qua việc kiểm tra bằng kính hiển vi của mình đối với các chế phẩm màng quan trọng trong suốt, nhà bệnh lý học người Đức Julius Cohnheim kết luận rằng phản ứng viêm về cơ bản là một hiện tượng mạch máu. Hiện tượng thực bào đã được mô tả vào cuối thế kỷ 19 bởi Elie Metchnikoff và các đồng nghiệp của ông tại Viện Pasteur. Các nghiên cứu hình thái học, sử dụng cả động vật sống và các chế phẩm mô học cố định, đã thay đổi hiểu biết của chúng ta về chứng viêm và dẫn đến các khái niệm hiện đang được nắm giữ về các thay đổi huyết động liên quan đến viêm, viêm cấp tính và viêm mãn tính. Trong suốt 5 thập kỷ qua, các kỹ thuật hiện đại về hóa sinh, nuôi cấy mô, sản xuất kháng thể đơn dòng, công nghệ DNA tái tổ hợp, và thao tác di truyền của các tế bào cô lập và toàn bộ động vật đã cho phép hiểu biết chi tiết hơn về các cơ chế phân tử và tế bào làm cơ sở cho phản ứng viêm, giải quyết tích cực tình trạng viêm, và chuyển sang sửa chữa và phục hồi mô.
1. Phản ứng viêm cấp là gì?
Viêm là phản ứng sinh lý ban đầu đối với tổn thương mô, chẳng hạn như phản ứng cơ học, nhiệt, điện, chiếu xạ, hóa chất hoặc nhiễm trùng. Phản ứng viêm cấp tính được đánh dấu nhất quán bởi sự rò rỉ vi mạch và sự tích tụ của bạch cầu trung tính. Bốn dấu hiệu cơ bản của viêm cấp tính, được đề cập ở trên, có thể được tính trong các thông số sinh lý của viêm. Các tác động toàn thân của tình trạng viêm, đặc biệt là viêm cấp tính, là nguyên nhân dẫn đến các dấu hiệu lâm sàng quen thuộc là sốt, phản ứng giai đoạn cấp tính và thay đổi cảm giác. Đến lượt mình, nhiễm trùng huyết là một hội chứng phản ứng viêm toàn thân xảy ra để phản ứng với nhiễm trùng; nhiễm trùng huyết nặng là nhiễm trùng huyết có biến chứng rối loạn chức năng cơ quan cấp tính; và sốc nhiễm trùng là tình trạng nhiễm trùng huyết phức tạp do hạ huyết áp, kháng thuốc hồi sức cấp dịch hoặc do tăng lactat huyết.
Tình trạng viêm cấp tính, bao gồm những thay đổi cục bộ trong lưu lượng máu, những thay đổi về tính thấm vi mạch và sự bài tiết bạch cầu trung tính. Ngoài những thay đổi về huyết động, tình trạng viêm bao gồm hoạt hóa tế bào nội mô, kết dính bạch cầu-nội mô ái lực thấp, tương tác kết dính bạch cầu-nội mô ái lực cao hoặc tĩnh tại, di chuyển bạch cầu, hoạt hóa bạch cầu, và làm giảm và phân giải phản ứng viêm sau đó. Sự di chuyển được điều chỉnh cao của bạch cầu từ hệ mạch vào các vị trí viêm và tế bào lympho qua các mô lympho thứ cấp và đến lượt nó, vào các vị trí vi sinh vật xâm nhập, là cốt lõi để bảo vệ vật chủ trong bối cảnh viêm nhiễm và miễn dịch. Sự phức tạp phi thường này của các quá trình điều tiết kiểm soát tình trạng viêm được minh chứng bởi, nhưng không giới hạn ở, các cytokine tiền viêm (ví dụ: yếu tố hoại tử khối u [TNF] - α , interleukin [IL] -1 β , IL-6) dẫn đến tình trạng viêm , chống lại sự gia tăng các cytokine chống viêm (ví dụ, IL-4, IL-10, IL-11, IL-13, yếu tố tăng trưởng biến đổi [TGF] - β , IL-1ra và các thụ thể cytokine hòa tan) làm giảm phản ứng viêm. Cả việc chấm dứt quá trình viêm và chuyển từ môi trường viêm tích cực sang môi trường tái tạo mô, chữa lành vết thương đều là những quá trình được điều chỉnh tích cực. Các chất trung gian này bao gồm các chất từ chất trung gian oxy và nitơ phản ứng tồn tại trong thời gian ngắn đến toàn bộ hệ thống điều hòa (ví dụ, bổ thể và đông máu). Nhiều tác nhân trung gian gây viêm đã trở thành mục tiêu cho các chiến lược điều trị gián đoạn.
2. Cơ chế của phản ứng viêm cấp
2.1. Thay đổi huyết động(Hemodynamic)
Những thay đổi huyết động xảy ra sớm trong viêm cấp tính bao gồm giãn mạch tiểu động mạch và tăng cục bộ tính thấm vi mạch. Trong nhiều trường hợp, giãn mạch tiểu động mạch theo sau một thời gian co mạch nhanh chóng và thoáng qua. Giãn mạch động mạch dẫn đến tăng lưu lượng máu, do đó giải thích hiện tượng sưng, nóng và đỏ quen thuộc đặc trưng cho vị trí viêm cấp tính. Sự gia tăng lưu lượng máu, cùng với sự gia tăng tính thấm vi mạch, dẫn đến sự cô đặc máu và tăng độ nhớt cục bộ. Những thay đổi huyết động học này rất quan trọng đối với sự di cư của bạch cầu tiếp theo vì tương tác kết dính nội mô-bạch cầu lăn qua trung gian selectin chỉ xảy ra trong những điều kiện lực cắt thấp như vậy. Tính thấm vi mạch tăng lên ban đầu dẫn đến sự dịch chuyển nghèo protein, sau đó là sự bài tiết huyết tương giàu protein, một đặc điểm khác của tình trạng viêm cấp tính.

Rò rỉ vi mạch xảy ra thông qua nhiều cơ chế được điều chỉnh theo thời gian, bao gồm sự co lại tế bào nội mô tĩnh mạch nhanh chóng, có thể đảo ngược và tồn tại trong thời gian ngắn do sự mở rộng các điểm nối gian bào; cái gọi là sự rút lại tế bào nội mô, ít được hiểu rõ hơn nhưng liên quan đến những thay đổi tế bào qua trung gian cytokine kéo dài; tổn thương nội mô trực tiếp và gián đoạn bởi chấn thương thực thể; tổn thương tế bào nội mô qua trung gian bạch cầu; và rò rỉ qua các mao mạch mới chưa có các điểm nối gian bào hoàn toàn “khép kín”. Tăng tốc độ chuyển tế bào mà các thành phần trong huyết tương đi qua các tế bào nội mô trong túi hoặc không bào (bào quan vesiculo vacuolar) xảy ra trong các mạch máu tân sinh và có thể đóng một vai trò trong viêm.
Sự thay đổi lưu lượng máu cục bộ xảy ra ở cấp độ tiểu động mạch, chìa khóa của sức cản mạch máu và được điều chỉnh phần lớn bởi hệ thống thần kinh tự chủ, nitric oxide (NO) (trước đây được gọi là yếu tố thư giãn có nguồn gốc từ nội mạc), peptide hoạt mạch và eicosanoids. Nhiều chất trung gian hóa học hòa tan có thể gây tăng tính thấm vi mạch thông qua một số cơ chế nêu trên.
2.2. Bạch cầu
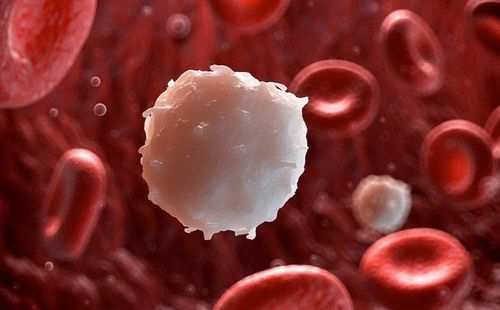
Việc tuyển chọn( huy động) bạch cầu vào vị trí viêm là đặc điểm cơ bản của phản ứng viêm. Việc tuyển dụng có tổ chức các loại bạch cầu cụ thể vào các mô cụ thể, cho dù là các vị trí viêm cấp tính, trong quá trình tái tuần hoàn tế bào lympho sinh lý qua các hạch bạch huyết hoặc trong phản ứng miễn dịch của tế bào đối với sự xâm nhập của vi sinh vật, được gọi là homing.
Các cơ chế chung của hoạt động di chuyển bạch cầu tương tự nhau, nhưng bạch cầu và các phân tử trung gian cụ thể khác nhau. Ví dụ, bạch cầu trung tính liên kết và đi ngang qua các tiểu tĩnh mạch sau mao mạch trong tình trạng viêm cấp tính, các tế bào lympho T chưa từng có liên kết và đi ngang qua các tiểu tĩnh mạch nội mô cao của hạch bạch huyết (HEV) trong tuần hoàn tế bào lympho, và tế bào lympho T có tác dụng và bộ nhớ liên kết và đi ngang qua các tế bào nội mô sau mao mạch ở các vị trí nhiễm trùng mãn tính.
Tầm quan trọng của các tế bào bạch cầu trong việc bảo vệ vật chủ được nhấn mạnh ở những bệnh nhân bị thiếu hụt số lượng bạch cầu hoặc khiếm khuyết chức năng. Bạch cầu rất quan trọng vì vai trò trung tâm của chúng trong quá trình thực bào và tiêu diệt hoặc ngăn chặn vi khuẩn và tiêu hóa các mảnh vụn mô hoại tử. Các sản phẩm có nguồn gốc từ bạch cầu, chẳng hạn như các enzym phân giải protein và các chất trung gian oxy phản ứng, góp phần làm tổn thương mô.
2.2.1. Sự kết dính và di chuyển bạch cầu
Sự ứ trệ mạch máu là kết quả của những thay đổi huyết động của tình trạng viêm cấp tính sớm dẫn đến sự di chuyển của bạch cầu từ cột trục trung tâm của các tế bào máu tuần hoàn đến các vị trí dọc theo bề mặt nội mô.
Quá trình này, hay sự kết hợp này( margination), được tăng cường trong điều kiện máu chảy chậm. Bạch cầu bám chắc và yếu vào bề mặt nội mô. Các chế phẩm màng sống và các nghiên cứu về buồng dòng chảy bằng cách sử dụng các đơn lớp tế bào nội mô và huyền phù của bạch cầu tinh khiết đã cho thấy rằng các tế bào “nhào lộn và lăn” dọc theo bề mặt nội mô. Tương tác kết dính nội mô bạch cầu trung tính nhẹ, yếu, thoáng qua xảy ra trong vòng vài phút sau khi bắt đầu phản ứng viêm cấp tính và có thể, tùy thuộc vào thời điểm trong quá trình tiến triển của phản ứng viêm, liên quan đến bạch cầu trung tính, tế bào lympho, bạch cầu đơn nhân, bạch cầu ưa bazơ hoặc bạch cầu ái toan. Tương tác kết dính cuốn tế bào nội mô-tế bào bạch cầu là một bước cụ thể và cần thiết trước khi có ái lực cao, hay còn gọi là kết dính và di chuyển tĩnh. Các tương tác kết dính lăn sớm được thực hiện chủ yếu nhờ các selectin và các chất phản ứng giàu carbohydrate của chúng. Đổi lại, sự biểu hiện trên bề mặt tế bào của selectin (và các phân tử kết dính gian bào khác) được điều chỉnh bởi các chất trung gian tiền viêm được sản xuất tại chỗ.

Selectin chứa vùng liên kết carbohydrate ở đầu N ngoại bào tương đồng với lectin của động vật có vú, vùng giống yếu tố tăng trưởng biểu bì, một loạt vùng điều hòa bổ thể và vùng xuyên màng ưa mỡ P-selectin được biểu hiện bởi các tế bào nội mô và tiểu cầu, E-selectin bởi các tế bào nội mô, và L-selectin bởi hầu hết các tế bào bạch cầu. P-selectin được tổng hợp thành phần và được lưu trữ trong các hạt nội chất trong tế bào chất (cơ thể Weibell-Palade).
Khi các tế bào nội mô tiếp xúc với histamine, thrombin, yếu tố hoạt hóa tiểu cầu (PAF) hoặc yếu tố mô, P-selectin đã được tạo sẵn sẽ nhanh chóng (trong vòng vài phút) được chuyển vị trí nhanh chóng (trong vòng vài phút) đến bề mặt nội mô, nơi nó tham gia vào các bạch cầu đã kết hôn thông qua các phân tử carbohydrate có chứa dư lượng axit sialic (ví dụ, P-selectin glycoprotein ligand-1 [PSGL-1]). Tương tác liên kết thoáng qua, có ái lực thấp này giải thích một phần cho các tương tác cuộn sớm. Sự phát triển của chuột loại trực tiếp (tức là thiếu selectin riêng lẻ [P - / -; E - / - hoặc L - / -], chuột loại trực tiếp đôi (ví dụ: E - / - và P - / -), và thậm chí là loại trực tiếp ba lần chuột, đã xác nhận rằng bạch cầu lăn có thể được gần như hoàn toàn chiếm bởi selectin. phơi sáng của các tế bào nội mô để TNF- α hoặc IL-1 β kết quả trong protein biểu hiện tổng hợp phụ thuộc mới của E-selectin, một phản ứng xảy ra trong vòng 1 đến 2 giờ và cao nhất là 4 đến 6 giờ.
Như trong trường hợp kết dính bạch cầu qua trung gian P-selectin, sự kết dính qua trung gian E-selectin xảy ra thông qua một loạt các gốc carbohydrate được sialyl hóa và fucosyl hóa liên quan đến các kháng nguyên nhóm máu sialyl Lewis X và sialyl Lewis A, nhưng được tìm thấy trên bạch cầu. Chất đối kháng Selectin bao gồm một số glycoprotein giống mucin khác nhau được bao phủ bởi các phần tử sialyl khác nhau. L-selectin được biểu hiện thành phần bởi bạch cầu, tham gia vào liên kết bạch cầu trung tính và bạch cầu đơn nhân với nội mô hoạt hóa ở các vị trí viêm, di chuyển tế bào lympho-tế bào nội mô (ví dụ, tế bào lympho di chuyển đến các hạch bạch huyết thông qua HEV), tương tác kết dính bạch cầu-bạch cầu thông qua mucin- chứa lưu huỳnh giống như glycoprotein, và được phân cắt khỏi tế bào nhờ các enzym “giảm” như disgrin và metalloproteinase (ADAM) -17 (enzym chuyển đổi TNF-α [TACE]) khi bạch cầu được kích hoạt.
Các thụ thể đối kháng glycoprotein giống mucin có liên quan trên HEV được gọi chung là địa chỉ nút ngoại vi (PNAds) và bao gồm phân tử bám dính tế bào địa chỉ niêm mạc (MadCAM) -1, GlyCAM-1 và CD34. L-selectin rụng đi tạo điều kiện thuận lợi cho sự di chuyển của bạch cầu bằng cách cho phép tế bào bạch cầu kết dính yếu tách ra khỏi nội mạc. Tương tác kết dính lăn có ái lực thấp tạo tiền đề cho các tương tác kết dính có ái lực cao qua trung gian β -và globulin miễn dịch và sự di chuyển bạch cầu.
Các tương tác cán mỏng qua trung gian selectin và chất kết dính tĩnh có ái lực cao không hoàn toàn rời rạc về mặt thời gian hoặc cơ học. Ví dụ, TNF- α và IL-1 β đều cảm ứng E-selectin, không được biểu hiện bởi các tế bào tĩnh lặng, và cả hai đều làm tăng biểu hiện nội mô của phân tử kết dính gian bào (ICAM) -1 và phân tử kết dính tế bào mạch (VCAM) -1, được thể hiện một cách cấu thành ở mật độ bề mặt tế bào thấp. ICAM-1 tham gia vào việc tuyển dụng tất cả các loại bạch cầu và VCAM-1 tham gia vào việc tuyển dụng các bạch cầu viêm mãn tính (tế bào lympho, bạch cầu đơn nhân, bạch cầu ái toan và basophils). ICAM-1 liên kết với Tích phân β 2 (bạch cầu), là cấu trúc dị số có chứa một loại chuỗi α (ví dụ: CD11a, CD11b, CD11c, CD11d) và chuỗi β chung (CD18). VCAM-1 liên kết với tích phân β 1 (ví dụ, kháng nguyên rất muộn [VLA] -4 / α 4 β 1 ) Các tế bào nội mô được kích hoạt tiết ra PAF và CXCL8 (IL-8), chúng hoạt hóa các bạch cầu liên kết với selectin bên trên.
Các dị phân tử CD11a / CD18 (LFA-1) của bạch cầu trải qua một sự thay đổi cấu trúc nhất thời và các nhóm phân tử CD11a / CD18 hình thành các cụm đa phân tử. Cả sự thay đổi cấu trúc trong CD11a / CD18 và sự phân nhóm đều góp phần làm tăng ái lực liên kết với ICAM-1 nội mô. CD11b / CD18 (Mac-1) liên kết với ICAM-1, ICAM-2 và iC3b (xem phần “Phần bổ sung”), phần sau của nó làm loại bỏ các hạt được phủ bổ thể. CD11c / CD18 cũng liên kết với iC3b và bắt đầu thực bào, nhưng đóng vai trò ít hơn trong việc kết dính bạch cầu trung tính so với CD11a / CD18 và CD11b / CD18. Các phân tử kết dính giữa các tế bào được tìm thấy trên nhiều loại tế bào khác nhau ngoài tế bào nội mô. Vai trò của CD11c / CD18, CD11d / CD18 và ICAM-3 trong kết dính bạch cầu-nội mô chưa được xác định rõ ràng. β 1 -Integrins, đặc biệt là VLA-4, được tìm thấy chủ yếu trên các bạch cầu viêm mãn tính (ví dụ, tế bào lympho, bạch cầu đơn nhân, basophils và bạch cầu ái toan) và làm trung gian gắn kết bạch cầu qua VCAM-1. β 1.Tương tác kết dính qua trung gian -integrin xảy ra thông qua chuỗi peptit axit arginine-glycine-aspartic (RGD) được hiển thị bởi VCAM-1, cũng như trên bề mặt tiếp xúc của các phân tử nền (ví dụ, fibronectin). Tương tác kết dính qua trung gian β 2 -Integrin – ICAM-1 và β 1 -integrin – VCAM-1 xảy ra muộn hơn (vài giờ đến vài ngày) trong phản ứng viêm so với tương tác qua trung gian selectin.
β 1 - và β 2 -integrins rõ ràng là rất quan trọng trong việc tuyển chọn bạch cầu vào các vị trí viêm cấp tính và mãn tính. Mười tám tiểu đơn vị tích phân α khác nhau , tám tiểu đơn vị tích phân β , và 24 tổ hợp heterodimer in vivo đã được xác định ở động vật có vú. Ngoài các tương tác kết dính tế bào nội mô-bạch cầu, các tích phân hoạt động trong một loạt các tương tác tế bào-tế bào và chất nền tế bào-ngoại bào khác. Một số lượng lớn các phân tử nhỏ, peptit và kháng thể thiết kế nhắm mục tiêu tích phân đã được phát triển cho các ứng dụng điều trị. Các bệnh về viêm - miễn dịch được điều trị bằng các chất đối kháng tích hợp bao gồm, trong số những bệnh khác, bệnh đa xơ cứng, bệnh Crohn và thoái hóa điểm vàng do tuổi tác.
Tương tác kết dính tĩnh tại có ái lực cao trước sự di chuyển của bạch cầu qua lớp nội mạc vào trung gian dưới da. Tầm quan trọng về mặt chức năng của tương tác kết dính giữa bạch cầu và nội mô đã được làm rõ bởi các nghiên cứu liên kết trong ống nghiệm và nghiên cứu in vivo đã sử dụng các kháng thể trung hòa chống lại các phân tử kết dính, chất đối kháng dược lý của các phân tử kết dính và chuột loại trực tiếp. Tầm quan trọng về mặt chức năng của tích phân bạch cầu (CD11a / CD18, CD11b / CD18, CD11c / CD18) cũng đã được nhấn mạnh bởi các quan sát lâm sàng và thực nghiệm ở những bệnh nhân mắc chứng thiếu kết dính bạch cầu di truyền hiếm gặp.
β 2 -Integrin ICAM-1 và ICAM-2, cũng như β 1 -integrin (VLA-4) VCAM-1, tạo ra các tương tác kết dính dẫn đến tổ chức lại tế bào trong bạch cầu làm phẳng và lan rộng trên bề mặt nội mô, mở rộng giả mô giữa các tế bào nội mô, và di chuyển dọc theo các gradient hóa học ngoại mạch. Hầu hết bạch cầu thoát ra khỏi không gian mạch máu giữa các tế bào nội mô liền kề (di cư nội bào). Sự di chuyển nội bào không chỉ phụ thuộc vào tương tác tích phân-phối tử mà còn phụ thuộc vào CD31 (PECAM-1 [phân tử kết dính tế bào nội mô tiểu cầu-1]) được biểu hiện trên cả bạch cầu và tế bào nội mô, và sự tháo rời thuận nghịch thoáng qua của nội mô mạch máu liên kết chặt chẽ (VE) - các phức chất liên kết cadherin. Có bằng chứng cho thấy bạch cầu cũng có thể thoát ra khỏi không gian mạch máu thông qua một con đường xuyên tế bào ít đặc trưng hơn.
2.2.2. Hóa chất và kích hoạt bạch cầu
Các bạch cầu liên kết chặt chẽ với nội mô di chuyển từ khoảng không gian mạch vào kẽ bằng cách kéo dài các nang giả giữa các điểm nối gian bào (Các protease trung tính được tiết ra như elastase, cathepsin G và proteinase 3, đóng một vai trò trong việc di chuyển hoặc “xâm nhập” của bạch cầu qua chất nền ngoại bào dưới nội mô. Collagenase đặc biệt quan trọng trong việc di chuyển bạch cầu qua màng đáy.
Nhiều loại metalloproteinase ma trận, được sản xuất bởi một số loại tế bào, tham gia vào quá trình di chuyển của bạch cầu, và cũng đóng vai trò trong việc giải quyết tình trạng viêm và tái tạo mô. Sự di chuyển của bạch cầu và sự di chuyển tiếp theo qua các kẽ theo các gradient nồng độ hóa chất; các quá trình được tạo điều kiện thuận lợi bằng các tương tác liên kết giữa các tích phân của bạch cầu và các vị trí bổ sung trên các phân tử nền ngoại bào (ví dụ, fibronectin). Nhiều chất hòa giải hòa tan có thể cung cấp động cơ thúc đẩy quá trình này.
Các yếu tố hóa học đối với bạch cầu trung tính bao gồm các peptit có nguồn gốc từ vi khuẩn (ví dụ, peptit N -formyl), peptide có nguồn gốc từ bổ thể (ví dụ, C5a), lipid hóa học có nguồn gốc từ màng tế bào (ví dụ, PAF), và các cytokine và chemokine được tạo ra bởi nhiều loại tế bào (ví dụ, CXCL8 [IL-8] từ tế bào nội mô). Các yếu tố hóa học thay đổi tùy theo tính đặc hiệu của chúng đối với các loại bạch cầu khác nhau. Ví dụ, C5a và N-formyl peptide đều gây ra phản ứng hóa học bạch cầu trung tính và bạch cầu đơn nhân, và CXCL8 [IL-8] gây ra điều hòa hóa học bạch cầu trung tính, trong khi CCL2 (protein hóa trị monocyte [MCP] -1) gây ra phản ứng hóa học trong bạch cầu đơn nhân và một tập hợp con cụ thể của tế bào lympho T bộ nhớ. Tập hợp con Th17 của tế bào lympho T trợ giúp CD4 tiết ra IL-17 và IL-22 tham gia vào quá trình tuyển dụng bạch cầu trung tính. (Có một số loài IL-17, bao gồm cả homodimers IL-17A – IL-17F cũng như các loài dị phân tử) Mỗi yếu tố hóa học này sẽ kích hoạt các tế bào “đích” bằng cách tham gia vào các thụ thể bề mặt tế bào cụ thể, do đó, được liên kết với bộ máy vận động co bóp của tế bào.
Ngoài tác dụng điều hòa hóa học, các chất trung gian hòa tan và bề mặt tế bào gây ra hoạt hóa bạch cầu được biểu hiện bằng một loạt các thay đổi trong chức năng tế bào (ví dụ, điều hòa tích phân bạch cầu và tăng ái lực liên kết [ví dụ, CD11b / CD18], rụng selectin [ví dụ, L-selectin ], phân giải lysosome và bắt đầu quá trình hô hấp). Đã có những tiến bộ lớn trong sự hiểu biết về các con đường sinh hóa liên quan đến quá trình điều hòa hóa học, hoạt hóa tế bào và quá trình suy giảm. Mặc dù có nhiều sắc thái trong các con đường dẫn truyền tín hiệu liên quan đến các quá trình này, một số chủ đề đã xuất hiện. Các thụ thể trên bề mặt tế bào được kích hoạt bởi các phối tử cụ thể (ví dụ: C5a, leukotriene B4 [LTB4], CXCL8 [IL-8] và sự hoạt hóa thụ thể được truyền tải thông qua các protein G cụ thể và các phospholipase liên kết màng, dẫn đến huy động canxi nội bào, dòng canxi ngoại bào và phosphoryl hóa hàng loạt protein tế bào. Bệnh di truyền hiếm gặp liên quan đến thụ và effector khuyết tật (ví dụ, IFN- γ cái nhìn sâu sắc dị tật bẩm thụ và adenine dinucleotide phosphate nicotinamide [giảm hình thức] [NADPH] oxidase khuyết tật) đã cung cấp vào chức năng bạch cầu và tầm quan trọng của hoạt động cụ thể như vậy để bảo vệ máy chủ.
Kết quả chính của việc tuyển dụng bạch cầu trung tính và bạch cầu đơn nhân là cung cấp một số lượng lớn bạch cầu hoạt hóa có thể giải phóng các chất lytic và các chất trung gian oxy và nitơ phản ứng cần thiết để tiêu diệt những kẻ xâm lược ngoại lai, và một phương tiện chứa các hạt lạ thông qua quá trình thực bào. Một số bạch cầu đơn nhân được tuyển chọn sẽ biệt hóa thành đại thực bào và các tế bào lympho có tác dụng và trí nhớ được tuyển dụng đóng vai trò quan trọng trong phản ứng miễn dịch thích ứng. Các sản phẩm và chức năng của các tế bào viêm đã được kích hoạt ngay lập tức được coi trọng vì chúng chứa và tiêu diệt những kẻ xâm lược và có hại vì chúng gây ra tổn thương mô. Vai trò của apoptosis bạch cầu trung tính (chết tế bào theo chương trình) trong việc chấm dứt các phản ứng viêm cấp tính và IFN- γ–Driven đại thực bào M1 và đại thực bào M2 điều khiển IL-4 / IL-13 trong quá trình chuyển đổi từ mô viêm sang mô chữa lành vết thương hoặc tái tạo mô được thảo luận trong “Đại thực bào M1 và M2”.
Sự hoạt hóa bạch cầu, đặc biệt là bạch cầu trung tính và thực bào đơn nhân, dẫn đến việc tiết ra các peptit diệt vi sinh vật (ví dụ, các chất khử trùng, protein làm tăng tính thấm diệt khuẩn [BPI], protein kháng khuẩn cation [ví dụ, CAP37]) và các enzym lytic (ví dụ: myeloperoxidase, elastase, cathepsin G). Việc giải phóng các thành phần dạng hạt này đi kèm với việc tạo ra các chất trung gian oxy và nitơ phản ứng (ví dụ, O 2 - , H 2 O 2 , NO), tạo ra các chất chuyển hóa arachidonat (ví dụ, leukotrienes và prostaglandin) và sản xuất các chất trung gian gây viêm khác. Trong một số trường hợp những chất trung gian được giải phóng vào phagolysosomes nơi nó đóng góp vào sự tàn phá của vi khuẩn nhấn chìm, trong khi ở các trường hợp khác nó được bài tiết vào môi trường ngoại bào nơi nó khuếch đại phản ứng và gây ra thiệt hại viêm mô. Các loại hạt bạch cầu trung tính khác nhau (azurophilic chính, đặc hiệu thứ cấp, chứa gelatinase bậc ba và các túi tiết) được giải phóng theo kiểu phối hợp khác nhau.
Để đặt lịch khám tại viện, Quý khách vui lòng bấm số HOTLINE hoặc đặt lịch trực tiếp TẠI ĐÂY. Tải và đặt lịch khám tự động trên ứng dụng MyVinmec để quản lý, theo dõi lịch và đặt hẹn mọi lúc mọi nơi ngay trên ứng dụng.