Tế bào mầm là loại tế bào duy nhất có khả năng tạo ra một sinh vật hoàn toàn mới. Để thực hiện các chức năng dành riêng cho dòng mầm và duy trì khả năng toàn năng, tế bào mầm ức chế sự phân hóa soma, tương tác với một môi trường vi mô chuyên biệt và sử dụng mạng lưới điều hòa RNA dành riêng cho mầm tế bào.
1. Tế bào mầm là gì?
Tế bào mầm là bất kỳ tế bào sinh học nào làm phát sinh giao tử của một sinh vật có khả năng sinh sản hữu tính. Ở nhiều loài động vật, tế bào mầm có nguồn gốc từ nguyên thủy và di chuyển qua ruột của phôi thai đến các tuyến sinh dục đang phát triển. Ở đó, chúng trải qua quá trình phân chia tế bào theo hai loại nguyên phân và meiosis, sau đó là sự phân hóa tế bào thành các giao tử trưởng thành, trứng hoặc tinh trùng. Không giống như động vật, thực vật không có tế bào mầm từ giai đoạn phát triển ban đầu. Thay vào đó, tế bào mầm có thể đến từ tế bào xôma ở người trưởng thành.
Trong hầu hết các sinh vật đa bào, tế bào mầm là nguồn gốc của các sinh vật mới cung cấp sự kế thừa thông tin di truyền và biểu sinh ở các thế hệ sau. Dòng tế bào mầm là nguồn gốc của sự toàn năng, cung cấp cho việc tạo ra các sinh vật mới. Vào thế kỷ 19, August Weismann đã công bố giả thuyết, theo đó, sự hiện diện của các yếu tố quyết định tế bào mầm đã hình thành sẵn (tế bào mầm) chỉ được di truyền thông qua các tế bào mầm đảm bảo tính toàn năng và tính liên tục của dòng mầm. Điều này đã được chứng minh ở động vật không xương sống và động vật có xương sống thấp hơn, nơi mà tế bào mầm cần thiết để hình thành tế bào mầm, trong khi ở động vật có vú, quá trình này liên quan đến cơ chế biểu sinh chứ không phải quá trình hình thành. Một số ảnh hưởng từ môi trường đóng một vai trò quan trọng và tính toàn năng được duy trì thông qua dòng mầm, cho phép phát triển hơn nữa ở thế hệ sau.
Tế bào mầm trải qua hai giai đoạn phát triển khác nhau đáng kể. Giai đoạn đầu tiên xảy ra trong quá trình hình thành phôi sớm, khi các tế bào mầm nguyên thủy (PGCs) được hình thành và tích cực di chuyển đến rãnh sinh dục. Trong giai đoạn thứ hai, các tế bào mầm nhận được các tín hiệu thích hợp từ môi trường của chúng và bắt đầu một trong hai chương trình phân chia tế bào có kiểm soát, quá trình meiosis, và biệt hóa - sinh trứng hoặc sinh tinh, để hình thành giao tử.

Cơ sở phân tử của cả hai quá trình và sự phát triển tế bào mầm ban đầu được hiểu rất rõ ở hai loài Drosophila và Caenorhabditis elegans, nơi các đánh giá di truyền có hệ thống đã xác định được nhiều gen cần thiết trong quá trình này. PGCs ở người chưa được nghiên cứu nhiều vì những trở ngại về kỹ thuật và đạo đức trong việc lấy các tế bào như vậy từ phôi ban đầu. Phần lớn kiến thức thu thập được về đặc điểm kỹ thuật PGC của động vật có vú thu được từ các nghiên cứu sử dụng phôi chuột ban đầu, trong đó, số tế bào mầm được tạo ra trong các tế bào biểu mô gần đa năng ngay sau khi được cấy vào thành tử cung.
2. Nguồn gốc và sự phát triển của tế bào mầm nguyên thủy
Ở động vật có vú, nguồn gốc của dòng tế bào mầm trong quá trình phát sinh phôi ban đầu không rõ ràng do không có dòng tế bào mầm đặc trưng có trong trứng như ở các sinh vật khác như X. laevis và D. melanogaster. Nguồn gốc và sự phát triển của tế bào mầm lần đầu tiên được xác định ở động vật có vú bởi Chiquoine vào năm 1954. Ông đã tìm thấy một quần thể các tế bào thuộc dòng tế bào mầm có khả năng tạo ra cả tế bào trứng và tinh trùng ở gốc của đồng phân mới nổi ở E7.25 trong nội bì của túi noãn hoàng của phôi chuột, ngay bên dưới vệt nguyên thủy, được xác định bởi phosphatase kiềm cao (AP) hoạt động.
Mặc dù những ràng buộc về đạo đức làm hạn chế những kiến thức của chúng ta về đặc điểm kỹ thuật của PGC ở người nhưng rõ ràng là các con đường tín hiệu chung hoạt động trên các loài động vật có vú và có thể là tất cả các động vật có xương sống. Dựa trên việc nhuộm AP, các nghiên cứu khác nhau đã gợi ý các vị trí nguồn gốc đa dạng cho PGC bao gồm cả vệt nguyên thủy phía sau. Quần thể sáng tạo của tế bào mầm có số lượng ít và sâu, điều này gây khó khăn lớn trong việc nghiên cứu cơ sở di truyền cho đặc điểm của dòng tế bào mầm. Các tế bào mầm, ngay sau khi hạn chế dòng dõi của chúng, có được hình thái phản ánh các đặc điểm phân tử cơ bản.
Saitou và cộng sự đã thiết lập một hệ thống để xác định các yếu tố chính quyết định số phận tế bào mầm ở chuột và để hiểu các đặc điểm khác biệt mà tế bào mầm có được ở cấp độ phân tử. Họ đã tách ra một vùng phôi với khoảng 300 tế bào có chứa tế bào mầm sáng lập ở E7.5 (giai đoạn EB) và phân tách nó thành các tế bào đơn lẻ. Các tế bào này giống nhau về mặt hình thái nhưng thuộc hai lớp có thể phân biệt được bằng sự biểu hiện khác biệt của hai gen đặc trưng cho tế bào mầm, Stella và Fragilis. Cụm 300 tế bào thể hiện sự biểu hiện phổ quát của Fragilis nhưng sự biểu hiện Stella bị giới hạn trong một tập hợp con các tế bào ở trung tâm của cụm. Do đó, cả hai gen dường như đều có vai trò chính trong sự phát triển tế bào mầm và khả năng biệt hóa của chúng. Stella là gen đầu tiên được biểu hiện trong quần thể tế bào được coi là tế bào mầm hạn chế dòng dõi cũng cho thấy sự biểu hiện cao của AP không đặc hiệu ở mô (Tnap), một gen cho hoạt động AP của PGC.

Tế bào mầm di cư tiếp tục biểu hiện rõ ràng và đặc hiệu của Stella nhưng lại biểu hiện sự đàn áp mạnh mẽ của tất cả các gen homeobox được khảo sát, mặc dù mức độ biểu hiện cao của các gen này ở các tế bào soma lân cận. Vì vai trò của gen homeobox là xác định danh tính khu vực của các tế bào dọc theo trục cơ thể hoặc tạo ra sự khác biệt của tế bào đối với các dòng tế bào soma cụ thể, điều này cho thấy rằng tế bào mầm có khả năng tránh đặc điểm soma bằng cách ngăn chặn hoặc ức chế sự biểu hiện của gen homeobox. Đây có thể là một trong những đặc điểm chính mà tế bào mầm của động vật có vú sở hữu cho phép chúng duy trì hoặc lấy lại sức mạnh và sự khác biệt với các tế bào xung quanh khác trong ngách. Khái niệm này được hỗ trợ bởi sự biểu hiện liên tục của Oct4 và các gen đa năng khác trong tế bào mầm.
3. Sự di cư của các tế bào mầm nguyên thủy
Trái ngược với D. melanogaster và cá ngựa vằn, người ta biết rất ít về sự di cư của PGC và bắt đầu quá trình đó ở chuột. Ngay sau khi đặc tả, các tế bào bắt đầu biểu hiện hình thái phân cực và sự mở rộng tế bào chất cũng như bắt đầu di chuyển qua vệt nguyên thủy vào nội bì phôi sau liền kề, nội bì ngoại phôi, và đồng phân. Bước đầu tiên trong quá trình di chuyển PGC của chuột là sự di chuyển của các tế bào từ vệt nguyên thủy sau đến nội bì tại E 7.5. Giữa E 8.5 và E 13.5, các PGC dương tính Tnap sinh sôi và di chuyển qua nội bì sau và mạc treo, di cư song phương đến các gờ sinh dục. Sau đó, chúng có thể xâm nhập vào meiosis ở con cái hoặc bắt đầu phân bào ở con đực và bắt đầu sự phân hóa thành tế bào trứng hoặc tinh trùng. Trong quá trình di cư, thời gian nhân đôi của “dân số” PGC khá đồng đều, vào khoảng 16 giờ từ 8,5 đến 13,5 ngày. Phôi chuột E 13.5 nên có khoảng 24000 PGC trong các gờ sinh dục của chúng. Trong giai đoạn di cư này, PGC trải qua quá trình tái lập trình rộng rãi bộ gen và thay đổi thông tin biểu sinh, chẳng hạn như sự methyl hóa DNA và các mẫu sửa đổi histone. Chúng cũng xóa dấu vết gen có thể cần thiết để khôi phục toàn năng cho dòng tế bào mầm.
Hiện tại, không có bằng chứng về sự khác biệt cụ thể về giới tính trong quá trình di cư của PGC ở bất kỳ sinh vật nào. Một tập hợp con các tế bào mầm trong tuyến sinh dục có được khả năng hoạt động như các tế bào gốc của dòng mầm, trải qua quá trình meiosis để tạo ra tinh trùng và trứng, đồng thời thúc đẩy thế hệ tiếp theo của sự phát triển phôi và di chuyển PGC. Protein Vasa là một thành phần thiết yếu của tế bào mầm và đại diện cho một phức hợp RNA và protein chưa được nghiên cứu rõ ràng cần thiết để xác định tế bào mầm. Đột biến rỗng dẫn đến bất dục ở chuột cái do những khiếm khuyết nghiêm trọng trong quá trình sinh sản.

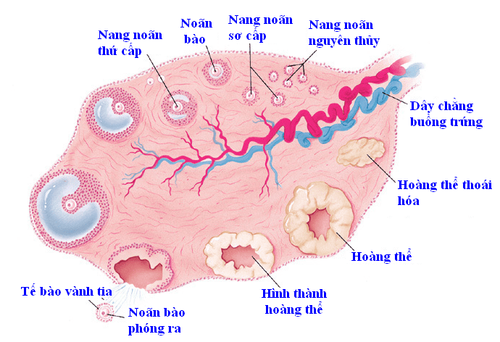
Ở người, sự biểu hiện VASA bắt đầu vào cuối giai đoạn di cư của quá trình phát triển PGC. Tilgner và cộng sự đã tạo ra và đặc trưng cho các dòng tế bào gốc phôi người (hESC) với cấu trúc trong đó sự biểu hiện của gen pEGFP-1 được điều khiển bởi trình tự DNA đại diện cho VASA report. Họ đã chứng minh rằng, hESC có thể được sử dụng như một hệ thống mô hình phát triển cho đặc điểm kỹ thuật PGC trong tương lai. Ngoài ra, họ có thể thiết lập một số lượng nhỏ các PGC nữ giả định được phân lập trên cơ sở phát huỳnh quang GFP do promoter VASA điều khiển. Những kết quả này và thực tế là sự biểu hiện cụ thể của Vasa trong dòng tế bào mầm trong quá trình thực dân của rãnh sinh dục cho thấy rằng Vasa là cần thiết để duy trì chức năng của tế bào mầm. Các PGC di cư duy trì một chương trình gen liên quan đến tính đa năng. Chúng biểu hiện các gen đa năng lõi (Oct4, Nanog và Sox2) và có khả năng hình thành u sau khi tiêm vào tinh hoàn của chuột sau sinh. Bên cạnh đó, các PGC di cư biểu hiện kháng nguyên phôi thai đặc hiệu giai đoạn 1 (SSEA1). Khi đến tuyến sinh dục, protein liên kết RNA đặc hiệu của tế bào mầm DAZL (bị xóa ở dạng azoospermia) là yếu tố cần thiết để phát triển PGC. Nhiều nghiên cứu cho thấy DAZL có chức năng như một chất tăng cường dịch mã dẫn đến những thay đổi nghiêm trọng về kiểu hình cho thấy DAZL có thể có các vai trò bổ sung trong giai đoạn PGC của quá trình phát sinh giao tử ở động vật có vú. DAZL cũng là tác nhân bảo vệ quá trình apoptosis ở PGC và điều chỉnh sự biểu hiện của các Caspase chính hoạt động như một cơ chế an toàn ngăn PGC lạc chỗ hình thành các khối u và loại bỏ PGC không ổn định. Trong trường hợp không có DAZL, các tế bào mầm không thể phát triển ngoài giai đoạn PGC như được thể hiện bằng cách tiếp tục biểu hiện các dấu hiệu đa năng. Những phát hiện này cho thấy DAZL là một “yếu tố cấp phép” cần thiết để phân biệt giới tính của PGC. Các PGC sau di cư, được đánh dấu bằng sự biểu hiện của một số protein liên kết RNA như MVH, DAZL và NANOS3, trải qua quá trình phát triển lưỡng hình hữu tính. Ở chuột, tế bào mầm cái nhanh chóng bắt đầu quá trình meiosis và bắt giữ ở giai đoạn meiosis I, trong khi những con đực phân chia nguyên phân trong vài vòng và sau đó bước vào giai đoạn tĩnh lặng khi chúng được gọi là tế bào sinh dục. Cụ thể, các PGC điều chỉnh một bộ gen cho phép chúng trải qua quá trình phân hóa giới tính và hình thành giao tử trong khi ngăn chặn sư hoạt động đa năng của chúng. Trong phôi giới nữ XX, các PGC tiếp tục tăng sinh tỷ lệ và sau đó tham gia vào giai đoạn giảm phân I. Sau khi sinh ra, các tế bào sinh dục được bao bọc bởi các tế bào từ lớp kẽ vỏ và trở thành tế bào trứng sơ cấp trong các nang nguyên thủy, từ đó chấm dứt tiềm năng tăng sinh tiền nhân và kìm hãm quá trình phát triển của chúng cho đến tuổi dậy thì. Sau tuổi dậy thì, sự kích thích nội tiết tố trong quá trình rụng trứng gây ra sự trưởng thành và phóng thích tế bào trứng từ buồng trứng vào ống dẫn trứng, tiếp theo là sự hoàn thành của phân chia meiotic đầu tiên đồng thời với sự phát triển của cơ thể. Khi thụ tinh với tế bào sinh tinh đơn bội, tế bào trứng hoàn thành lần phân chia cộng sinh thứ hai và tạo ra thể cực thứ hai. Ngược lại với nữ, PGC của nam giới XY bắt đầu phân bào khi xâm nhập vào các gờ sinh dục và ở trạng thái tĩnh lặng trong giai đoạn của chu kỳ tế bào trong thời kỳ phôi thai còn lại như một phôi thai, trong khi vẫn giữ được tiềm năng tiền sinh sản. Vào khoảng ngày thứ 5 sau khi sinh, nhiều pro spermatogonia tiếp tục tăng sinh tích cực, trong khi một số di chuyển đến màng đáy của ống bán lá kim và hình thành các điểm nối chặt chẽ với các tế bào Sertoli, do đó hình thành các tế bào gốc sinh tinh (SSC) được kết hợp vào ngách thích hợp của chúng. So với kích thước rất hạn chế của bể tế bào trứng, tế bào tinh trùng có thể thu được từ sự biệt hóa của SSC. Trong nuôi cấy, tế bào gốc dòng mầm có khả năng tăng sinh lâu dài và sinh tinh khi cấy ghép vào tinh hoàn được thiết lập với sự hiện diện của GDNF (yếu tố dinh dưỡng thần kinh có nguồn gốc từ tế bào thần kinh đệm), dễ có nhất từ tinh hoàn của trẻ sơ sinh.Khả năng tạo ra các giao tử trưởng thành từ PGC đại diện cho một lĩnh vực nghiên cứu cung cấp cái nhìn sâu sắc hơn về các con đường tín hiệu của quá trình phát sinh giao tử và rối loạn chức năng sinh sản ở người. Những tiến bộ rất gần đây trong công việc phân biệt có mục tiêu của iPSC cung cấp các liệu pháp iPSC được thiết kế riêng / cá nhân hóa cũng để điều trị chứng azoospermia ở nam giới hoặc suy buồng trứng nguyên phát ở nữ giới. Tuy nhiên, các vấn đề về nguồn gốc của tế bào tinh trùng và tế bào trứng trong điều kiện in vitro ở người vẫn còn: Sự biệt hóa của tế bào mầm phụ thuộc vào môi trường soma hơn là hàm lượng nhiễm sắc thể giới tính của tế bào mầm. Do đó, việc chứng minh sự phát sinh giao tử từ chuột ESC và iPSC cho thấy rằng các giao tử “nhân tạo” của con người cũng có thể thu được theo cách này và sự ra đời trực tiếp của chuột con cho thấy rằng các tế bào của con người cũng có thể hoạt động. Mặc dù phương pháp này đang rất hứa hẹn nhưng nó cũng đặt ra nhiều vấn đề về mặt đạo đức. Tuy nhiên, việc thu được tinh trùng và tế bào trứng còn sống của con người trong điều kiện trong phòng thí nghiệm chắc chắn sẽ mở ra cho y học sinh sản một hình thức điều trị vô sinh mới.
Để đặt lịch khám tại viện, Quý khách vui lòng bấm số HOTLINE hoặc đặt lịch trực tiếp TẠI ĐÂY. Tải và đặt lịch khám tự động trên ứng dụng MyVinmec để quản lý, theo dõi lịch và đặt hẹn mọi lúc mọi nơi ngay trên ứng dụng.
Nguồn tham khảo: cell.com, hindawi.com